|
||||||||||||||
Back to Parasitoids Table of Contents
Pseudacteon spp. by Lloyd W. Morrison, Department of Biology, Missouri State University, 901 S National Ave, Springfield, MO 65897 Phorid flies in the genus Pseudacteon are parasitoids of ants. Several ant genera serve as hosts, including Crematogaster, Lasius, Linepithema, and Solenopsis (Disney 1994). Here I focus on Pseudacteon species that parasitize Solenopsis fire ants, as this is the group on which most research has been done. Over 20 Pseudacteon species are known to parasitize Solenopsis saevissima complex fire ants in South America (Patrock et al. 2009). Similarly, more than 20 Pseudacteon species parasitize Solenopsis geminata complex fire ants from North America to northern South America (Plowes et al. 2009). None of the North or Central American Pseudacteon species parasitize the red imported fire ant, S. invicta, or the black imported fire ant, S. richteri, both accidentally introduced from South America and now major pests in the United States. Pseudacteon species from South America that parasitize S. saevissima complex ants represent classical biological control agents for imported Solenopsis fire ants in the United States. The biology and natural history of Pseudacteon phorids that parasitize Solenopsis fire ants have been summarized by Porter (1998a) and Morrison (2000a).
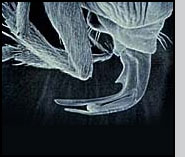
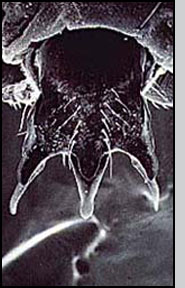
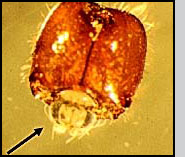
Appearance Pseudacteon phorid flies are very small, about the size of their host ant's head. Seen under a microscope, adult Pseudacteon flies have relatively large eyes, a sort of humped back ('humpbacked' flies are one of the common names for this family) and in females, an elaborately shaped ovipositor that varies among species. In the field, they appear as minute, fuzzy specks as they hover over host ants.
Habitat Pseudacteon species that parasitize Solenopsis fire ants are widely distributed in the natural range of their hosts. In tropical and subtropical areas, Pseudacteon species are active all year (Folgarait et al. 2003, Morrison and Porter 2005a, Calcaterra et al. 2008). In temperate regions, Pseudacteon adults are active in all except the winter months (Morrison et al. 1999a). Pseudacteon phorids are not known to be attracted in large numbers to anything other than host ants. Many phorids appear to be generalists, feeding on honeydew, plant sap, nectar, and dead insects, and Pseudacteon adults are also likely to be generalist feeders.
Pests Attacked (Host Range) Individual Pseudacteon spp. are almost always restricted to a single ant genus (Disney 1994). A given Pseudacteon species may attack multiple Solenopsis spp., but is usually restricted to a single species complex, and often reveals affinities for a single species. The host specificity of Pseudacteon that attack Solenopsis has been documented in tests conducted: (1) in the field in South America (Porter et al. 1995a, Porter 1998b), (2) in the lab prior to the release of South American Pseudacteon species to North America (Gilbert and Morrison 1997, Porter and Alonso 1999), and (3) in the field in North America after the establishment of introduced populations (Vazquez and Porter 2005, Morrison and Porter 2006).
Life Cycle Pseudacteon are solitary parasitoids. Each female Pseudacteon may produce 100 - 300 eggs (Zacaro and Porter 2003). The adult female inserts a single egg into a worker ant with a hypodermic-style ovipositor in a rapid aerial attack. The egg is inserted into the thorax region, and the larva migrates to the head capsule of the worker. There are three instars, during which time worker ants appear to behave normally until just before pupariation.
Relative Effectiveness Pseudacteon parasitism rates have been documented to be very low (<3%) (Morrison et al. 1997, Morrison and Porter 2005a, Calcaterra et al. 2008). In the presence of Pseudacteon species, however, Solenopsis worker behavior is affected and foraging is disrupted, leading to dramatic short-term decreases in resource retrieval (Feener and Brown 1992, Porter et al. 1995c, Orr et al. 1995, Folgarait and Gilbert 1999, Morrison 1999, 2000b). Although forager numbers may decrease, Solenopsis workers do not usually completely abandon food resources in the presence of Pseudacteon, but some workers remain behind to guard the resource (Porter et al. 1995c, Orr et al. 1997, Morrison et al. 2000). Host Solenopsis workers have sometimes been observed to lose control of food resources to competing ant species in the presence of Pseudacteon flies (Orr et al. 1995, Feener et al. 2008), although most of the time they appear to remain in control of the resource (Morrison 1999, Morrison et al. 2000, Feener et al. 2008). It is unclear to what degree the observed short-term behavioral effects may translate into long-term population-level impacts. A multi-year field experiment in Florida did not find any measurable effect of introduced P. tricuspis on S. invicta abundance (Morrison and Porter 2005b). Conservation Specific information on how to conserve Pseudacteon is not known. For general information about conservation of natural enemies, see Conservation in the Tutorial section on this site, or the Volume II, No. 1 Feature Article on conservation in the Midwest Biological Control News Online.
Pesticide Susceptibility Little is known. It has been found that exposure of parasitized ants to methoprene and pyriproxyfen—insect growth regulators that are the active ingredient in some fire ant baits—can reduce P. tricuspis emergence (Farnum and Loftin 2010).
Commercial Availability Pseudacteon species are not commercially available. However, selected Pseudacteon species are being mass-reared and released in many southeastern states. It is estimated that P. tricuspis now occurs in 65%, while P. curvatus may occur in as much as 90%, of the invasive S. invicta/S. richteri range in the southeastern US (Callcott et al. 2011). Two other species—P. litoralis and P. obtusus—have been established in localized areas, and releases of additional species are planned (Callcott et al. 2011).
References Calcaterra, L. A., A. Delgado and N. D. Tsutsui. 2008. Activity patterns and parasitism rates of fire ant-decapitating flies (Diptera: Phoridae: Pseudacteon spp.) in their native Argentina. Annals of the Entomological Society of America 101: 539-550. Callcott, A.-M., S. D. Porter, R. D. Weeks, Jr., L. C. Graham, S. J. Johnson and L. E. Gilbert. 2011. Fire ant decapitating fly cooperative release programs (1994-2008): Two Pseudacteon species, P. tricuspis and P. curvatus, rapidly expand across imported fire ant populations in the southeastern United States. Journal of Insect Science 11:19 available online: insectscience.org/11.19 Cônsoli, F. L., C. T. Wuellner, S. B. Vinson and L. E. Gilbert. 2001. Immature development of Pseudacteon tricuspis (Diptera: Phoridae), an endoparasitoid of the red imported fire ant (Hymenoptera: Formicidae). Annals of the Entomological Society of America 94: 97-109. Disney, R. H. L. 1994. Scuttle Flies: The Phoridae. Chapman & Hall, London. Farnum, J. M. and K. M. Loftin. 2010. Impact of methoprene and pyriproxyfen on Pseudacteon tricuspis (Diptera: Phoridae), a parasitoid of the red imported fire ant, Solenopsis invicta (Hymenoptera: Formicidae). Florida Entomologist 93: 584-589. Feener, D. H., Jr. and B. V. Brown. 1992. Reduced foraging of Solenopsis geminata (Hymenoptera: Formicidae) in the presence of parasitic Pseudacteon spp. (Diptera: Phoridae). Annals of the Entomological Society of America 85: 80-84. Feener, D. H., Jr., M. R. Orr, K. M. Wackford, J. M. Longo, W. W. Benson and L. E. Gilbert. 2008. Geographic variation in resource dominance-discovery in Brazilian ant communities. Ecology 89: 1824-1836. Folgarait, P. J., O. A. Bruzzone and L. E. Gilbert. 2003. Seasonal patterns of activity among species of black fire ant parasitoid flies (Pseudacteon: Phoridae) in Argentina explained by analysis of climatic variables. Biological Control 28: 368-378. Folgarait, P. J. and L. E. Gilbert. 1999. Phorid parasitoids affect foraging activity of Solenopsis richteri under different availability of food in Argentina. Ecological Entomology 24: 1663-173. Gilbert, L. E. and L. W. Morrison. 1997. Patterns of host specificity in Pseudacteon parasitoid flies (Diptera, Phoridae) that attack Solenopsis fire ants (Hymenoptera, Formicidae). Environmental Entomology 26: 1149-1154. Morrison, L. W., C. G. Dall'Aglio-Holvorcem and L. E. Gilbert. 1997. Oviposition behavior and development of Pseudacteon flies (Diptera: Phoridae), parasitoids of Solenopsis fire ants (Hymenoptera: Formicidae). Environmental Entomology 26: 716-724. Morrison, L. W. and L. E. Gilbert. 1998. Parasitoid-host relationships when host size varies: the case of Pseudacteon flies and Solenopsis fire ants. Ecological Entomology 23: 409-416. Morrison, L. W. 1999. Indirect effects of phorid fly parasitoids on interspecific competition between fire ants. Oecologia 121: 113-122. Morrison, L. W., E. A. Kawazoe, R. Guerra and L. E. Gilbert. 1999a. Phenology and dispersal in Pseudacteon flies (Diptera: Phoridae), parasitoids of Solenopsis fire ants (Hymenoptera: Formicidae). Annals of the Entomological Society of America 92: 198-207. Morrison, L. W., S. D. Porter, and L. E. Gilbert. 1999b. Sex ratio variation as a function of host size in Pseudacteon flies (Diptera: Phoridae), parasitoids of Solenopsis fire ants (Hymenoptera: Formicidae). Biological Journal of the Linnean Society 66: 257-267. Morrison, L. W. 2000a. Biology of Pseudacteon (Diptera: Phoridae) ant parasitoids and their potential to control imported Solenopsis fire ants (Hymenoptera: Formicidae). In S. G. Pandalai (ed.), Recent Research Developments in Entomology 3: 1-13. Morrison, L. W. 2000b. Mechanisms of Pseudacteon parasitoid (Diptera: Phoridae) effects on exploitative and interference competition in host Solenopsis ants (Hymenoptera: Formicidae). Annals of the Entomological Society of America 93: 841-849. Morrison, L. W., E. A. Kawazoe, R. Guerra and L. E. Gilbert. 2000. Ecological interactions of Pseudacteon parasitoids and Solenopsis ant hosts: environmental correlates of activity and effects on competitive hierarchies. Ecological Entomology 25: 433-444. Morrison, L. W. and J. R. King. 2004. Host location behavior in a parasitoid of imported fire ants. Journal of Insect Behavior 17: 367-383. Morrison, L. W. and S. D. Porter. 2005a. Phenology and parasitism rates in introduced populations of Pseudacteon tricuspis, a parasitoid of Solenopsis invicta. BioControl 50: 127-141. Morrison, L. W. and S. D. Porter. 2005b. Testing for population-level impacts of introduced Pseudacteon tricuspis flies, phorid parasitoids of Solenopsis invicta fire ants. Biological Control 33: 9-19. Morrison, L. W. and S. D. Porter. 2006. Post-release host-specificity testing of Pseudacteon tricuspis, a phorid parasitoid of Solenopsis invicta fire ants. BioControl 51: 195-205. Orr, M. R., S. H. Seike, W. W. Benson and L. E. Gilbert. 1995. Flies suppress fire ants. Nature 373: 292. Orr, M. R., S. H. Seike and L. E. Gilbert. 1997. Foraging ecology and patterns of diversification in dipteran parasitoids of fire ants in south Brazil. Ecological Entomology 22: 305-314. Patrock, R, W., S. D. Porter, L. E. Gilbert, and P. J. Folgarait. 2009. Distributional patterns of Pseudacteon associated with the Solenopsis saevissima complex in South America. 17pp. Journal of Insect Science 9:60, available online: insectscience.org/9.60 Plowes, R. M., E. G. LeBrun, B. V. Brown and L. E. Gilbert. 2009. A review of Pseudacteon (Diptera: Phoridae) that parasitize ants of the Solenopsis geminata complex (Hymenoptera: Formicidae). Annals of the Entomological Society of America 102: 937-958. Porter, S. D., H. G. Fowler, S. Campiolo, and M. A. Pesquero. 1995a. Host specificity of several Pseudacteon parasites of fire ants in South America (Diptera: Phoridae; Hymenoptera: Formicidae). Florida Entomologist 78: 70-75. Porter, S. D., M. A. Pesquero, S. Campiolo and H. G. Fowler. 1995b. Growth and development of Pseudacteon phorid fly maggots (Diptera: Phoridae) in the heads of Solenopsis fire ant workers (Hymenoptera: Formicidae). Environmental Entomology 24: 475-479. Porter, S. D. 1998a. Biology and behavior of Pseudacteon decapitating flies (Diptera: Phoridae) that parasitize Solenopsis fire ants (Hymenoptera: Formicidae). Florida Entomologist 81: 292-309. Porter, S. D. 1998b. Host-specific attraction of Pseudacteon flies (Diptera: Phoridae) to fire ant colonies in Brazil. Florida Entomologist 81: 423-429 Porter, S. D. and L. E. Alonso. 1999. Host specificity of fire ant decapitating flies (Diptera: Phoridae) in laboratory oviposition tests. Journal of Economic Entomology 92: 110-114. Vazquez, R. J. and S. D. Porter. 2005. Re-confirming host specificity of the fire ant decapitating fly Pseudacteon curvatus (Diptera: Phoridae) after field release in Florida. Florida Entomologist 88: 107-110. Zacaro, A. A., and S. D. Porter. 2003. Female reproductive system of the decapitating fly Pseudacteon wasmanni Schmitz (Diptera: Phoridae). Arthropod Structure and Development 31:329-337.
Back to Parasitoids Table of Contents
|
|
|||||||||||||