|
||||||||||||
Back to Parasitoids Table of Contents
Catolaccus grandis (Burks) by Juan A. Morales-Ramos, USDA-ARS, Southern Regional Research Center, New Orleans, LA 70124 Catolaccus grandis is an ectoparasitoid of the boll weevil classified initially within the genus Heterolaccus (Burks 1954). It was first introduced to the U.S. during the early 1970’s and released in experimental fields in Mississippi State, MS (Johnson et al. 1973). Researchers observed encouragingly high parasitism rates, but failure of this parasitoid to become established made it unsuitable for use within a classical biological control program. It was reintroduced a decade later and released in College Station, TX (Cate et al. 1990). Rearing methods for this parasitoid progressed quickly between 1985 and 1992 making C. grandis suitable to be used in a biological control by augmentation program (Cate 1987, Morales-Ramos et al. 1994).
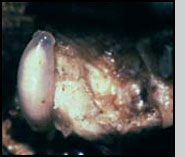
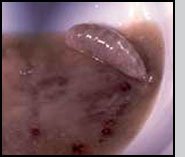
Appearance The female has a length between 4 to 5.5 mm. The head and thorax are shining black with the antennal pedicel and scape yellow or light tan, the funicle and club brown. The compound eyes are bright red. The legs have black coxa shading to tan, the trocanters and femora are tan and the tibia and basal tarsal segments are light yellow. The abdomen is red-brown with iridescent (metallic) blue or blue-green reflections. Males are smaller, 3.0-3.5 mm in length, and their abdomen differs in shape and coloration from that of females. The female abdomen is pointed metallic blue or blue-green, while the male abdomen is oval in shape and presents a transversal window white to light yellow in coloration. Male and female C. grandis during mating. Note ovoid male abdomen compared to pointed female abdomen and the size difference between the sexes. Photo: J. Morales-Ramos
Habitat Catolaccus grandis originated in Southeastern Mexico and it occurs naturally in the Mexican states of Veracruz, Tabasco, Campeche, Yucatan, Chiapas, Nayarit and Sinaloa (Cross and Mitchell 1969, Cate et al. 1990). This parasitoid lives in the tropical and subtropical forest and parasitizes its natural host (Anthonomus grandis) in native host plants, which include Hampea nutricia Fryxell, H. trilobata Standley, Cienfuegosia rosei Fryxel, and also in wild and cultivated cotton, Gossypium hirsutum L. (Cate et al. 1990). Catolaccus grandis is a tropical insect which is not adapted to overwinter in temperate regions. In the states of Tabasco, Campeche, Veracruz, and Chiapas, this parasitoid inhabits the rain forest in clearings where the Hampea plants usually grow. Dryer climates are also suitable for this parasitoid. In the states of Yucatan, Nayarit, Chiapas, and Sinaloa, this parasitoid survives in wild or cultivated cotton and short brush where Cienfuegosia plants proliferate.
Pests Attacked Only two species are known to be natural hosts of C. grandis: the boll weevil, Anthonomus grandis and a closely related species, Anthonomus hunteri, which inhabits the Yucatan peninsula. This parasitoid seems to be specific and highly adapted to its natural host; however, under laboratory conditions C. grandis has been successfully reared in three other species including the cow pea weevil, Callosobruchus maculatus (F.), the cow pea curculio Chalcodermus aeneus Boheman, and the pepper weevil Anthonomus eugenii Cano with significant reduction in fitness (Rojas et al. 1998, 1999). Nevertheless, C. grandis is so specific that it is unlikely to have an effect on insect species other than the boll weevil.
Life Cycle Catolaccus grandis is an idiobiont parasitoid (Quicke 1997), e.g., it arrests host development upon parasitism. Some of its characteristics relating to this trait include:
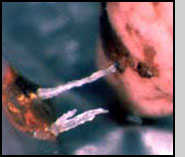
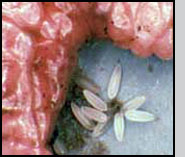
Females can locate cotton buds (squares) infested by boll weevils at ranges up to 60 m (Morales-Ramos and King 1991, Coleman et al. 1995). Females probe infested squares many times from different angles with their ovipositor. If the square contains a suitable host, the females paralyze the weevil larva and oviposit one single egg inside the cavity next to the paralyzed weevil (Morales-Ramos et al. 1995a). The preferred host stages for ovipositing are third instar and early pupa (Morales-Ramos and Cate 1992a). Females may or may not feed on the host by constructing a feeding tube (Morales-Ramos et al. 1996). The egg hatches in 18-24 h depending on the temperature. The first instar parasitoid is very active and moves around the cavity until it finds the host, then it attaches externally to feed on it. The larval stage consists of 5 stadia and lasts between 6 to 9 days, the pupal stage lasts between 7 to 9 days (Morales-Ramos and Cate 1993). The adults emerge and cut a circular exit hole in the square. The sex ratio is approximately 3 to 4 females per male. Mating occurs within an hour of emergence and females mate only once (Morales-Ramos and Cate 1992a). Females have a preovipositional period of 2 to 5 days during which they are not interested in host seeking.
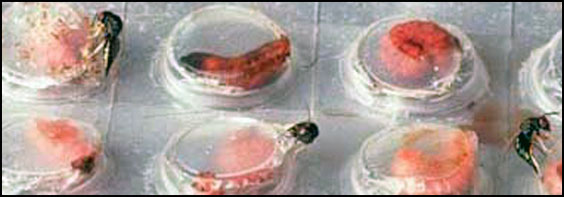
Relative Effectiveness Augmentative releases of C. grandis are very effective against boll weevil infestations. Parasitism rates of 70 to 90% have been reported consistently (King et al. 1995, Morales-Ramos et al. 1994, 1995b, Summy et al. 1994a, 1995a, 1995b, Vargas-Complis et al. 1997). The high searching capacity and specificity of this parasitoid makes it ideal for use in inundative releases. Weekly releases of 500 females per acre for 6 consecutive weeks are sufficient to achieve excellent control of the boll weevil, eliminating economical damage by this pest (Coleman et al. 1996, King et al. 1995, Summy et al. 1995a). However, C. grandis must be released every year because it does not overwinter in the U.S. The main obstacle for commercial application of C. grandis is the high cost and complexity required for its mass propagation. An artificial diet has been developed to rear C. grandis (Rojas et al. 1996), but a small quantity of host larvae are still required for the conditioning of adult females (Morales-Ramos et al. 1996). Parasitoids produced in vitro have been tested in laboratory and field and they have performed satisfactorily (Morales-Ramos et al. 1998).
Photo: J. Morales-Ramos
Conservation After inundative release of C. grandis, this parasitoid remains in the field as long as there are hosts available for reproduction. The use of nectaried (with extrafloral nectaries) cotton varieties enhances the survival and fecundity of adult parasitoids (Morales-Ramos and Cate 1992b). The use of mechanical cultivation reduces parasitism rates by C. grandis because burying infested squares protects boll weevils from parasitism (Summy et al. 1994b). Limiting cultivation practices to the beginning of the cotton-growing season reduces its impact on the parasitoids.
Pesticide Susceptibility As with most parasitoid wasps, C. grandis is often more susceptible to insecticides than its host. This parasitoid is highly susceptible to organophosphates such as methyl parathion and malathion. The toxicity to C. grandis adults of 10 insecticides including dimethoate, endosulfan, oxamyl, acephate, malathion, azinphos-methyl, cyfluthrin, methyl parathion, spinosad, and fipronil were tested. All of them were highly toxic to the parasitoids although endosulfan showed the least toxicity (Elzen 1998). Methyl parathion was the most toxic to C. grandis (Elzen 1998), but its low residual effect makes it useful for early applications (prior to releases) against fleahoppers and overwintering weevils (Summy et al. 1994a). Insecticides that remain toxic in the field for more than a week, such as oxamyl, thiodicarb, and azinphosmethyl were detrimental to the effectiveness of inundative releases (Summy et al. 1994a).
Commercial Availability Catolaccus grandis is not commercially available yet. This parasitoid was mass-produced for experimental releases at the USDA-ARS Subtropical Agricultural Research Center in Weslaco, Texas from 1990 to 2000. New technology to facilitate commercial scale production of this parasitoid was under development (Edwards et al. 1998), but the ongoing Boll Weevil Eradication Program precludes further efforts at this time.
References Burks, B. D. 1954. Parasitic wasps of the Catolaccus group in the Americas. USDA, Technical Bull. No. 1093: 1-21. Cate, J. R. 1987. A method of rearing parasitoids of boll weevil without the host plant. Southwest. Entomol. 12: 211-215. Cate, J. R., P. C. Krauter and K. E. Godfrey. 1990. Pests of cotton, pp. 17-29. En D. H. Habeck, F. D. Bennett, and J. H. Frank, Classical Biological Control in the Southern United States. South. Coop. Ser. Bull. 355. 197 pp. Coleman, R. J., S. M. Greenberg and E. G. King. 1995. Dispersal of female Catolaccus grandis in cotton, pp. 34-36. In Addendum to the Proceedings Beltwide Cotton Conferences 1995, National Cotton Council of America, Memphis, TN. Coleman, R. J., J. A. Morales-Ramos, E. G. King, and L. A. Wood. 1996. Suppression of the boll weevil in organic cotton by release of Catolaccus grandis as part of the Southern Rolling Plain boll weevil eradication program, p. 1094. In Proceedings Beltwide Cotton Conferences 1996, Vol. 2, National Cotton Council of America, Memphis, TN. Cross, W. H. and H. C. Mitchell. 1969. Distribution and importance of Heterolaccus grandis as a parasite of the boll weevil. Ann. Entomol. Soc. Am. 62: 235-236. Edwards, R. H., J. A. Morales-Ramos, M. G. Rojas, and E. G. King. 1998. Proposed transformation of laboratory to commercial-scale mass rearing of Catolaccus grandis (Hymenoptera: Pteromalidae). Vedalia 5: 133-139. Elzen, G. W., M. G. Rojas, P. J. Elzen, E. G. King, and N. M. Barcenas. 1998. Toxicity of insecticides to the boll weevil parasitoid Catolaccus grandis (Hymenoptera: Pteromalidae). Vedalia 5: 123-131. Godfray, H. C. J. 1994. Parasitoids, Behavioral and Evolutionary Ecology. Princeton University Press. Princeton, MA. Heinz, K. M. 1998. Host size-dependent sex allocation behavior in a parasitoid: implications for Catolaccus grandis (Hymenoptera: Pteromalidae) mass rearing programmes. Bull. Entomol. Res. 88: 37-45. Johnson, W. L., W. H. Cross, W. L. McGovern and H. C. Mitchell. 1973. Biology of Heterolaccus grandis in a laboratory culture and its potential as an introduced parasite of the boll weevil in The United States. Environ. Entomol. 2: 112-118. King, E. G., R. J. Coleman, L. Woods, L. Wendel, S. Greenberg, and A. W. Scott. 1995. Suppression of the boll weevil in commercial cotton by augmentative releases of a wasp parasite, Catolaccus grandis, pp. 26-30. In Addendum to the Proceedings Beltwide Cotton Conferences 1995, National Cotton Council of America, Memphis, TN. Morales-Ramos, J. A. and E. G. King. 1991. Evaluation of Catolaccus grandis (Burks) as a biological control agent against the cotton boll weevil, p. 724. In Proceedings Beltwide Cotton Conferences 1991, Vol. 2, National Cotton Council of America, Memphis, TN. Morales-Ramos, J. A. and J. R. Cate. 1992a. Laboratory determination of age-dependent fecundity, development, and rate of increase of Catolaccus grandis (Burks) (Hymenoptera: Pteromalidae). Ann. Entomol. Soc. Am. 85: 469-476. Morales-Ramos, J. A. and J. R. Cate. 1992b. Functional response of Catolaccus grandis (Burks) (Hymenoptera: Pteromalidae) in field cages. Biol. Cont. 2: 193-202. Morales-Ramos, J. A. and J. R. Cate. 1993. Temperature-dependent developmental rates of Catolaccus grandis (Burks) (Hymenoptera: Pteromalidae). Environ. Entomol. 22: 226-233. Morales-Ramos, J. A., K. R. Summy, J. L. Roberson, J. R. Cate, and E. G. King. 1992. Feasibility of mass rearing of Catolaccus grandis, a parasitoid of the boll weevil, pp. 723-726. In Proceedings Beltwide Cotton Conferences 1992, Vol. 2, National Cotton Council of America, Memphis, TN. Morales-Ramos, J. A., M. G. Rojas, J. Roberson, R. G. Jones, E. G. King, K. R. Summy and J. R. Brazzel. 1994. Suppression of the boll weevil first generation by augmentative releases of Catolaccus grandis in Aliceville, Alabama, pp. 958-964. In Proceedings Beltwide Cotton Conferences 1994, Vol.2, National Cotton Council of America, Memphis, TN. Morales-Ramos, J. A., M. G. Rojas and E. G. King. 1995a. Venom of Catolaccus grandis (Hymenoptera: Pteromalidae) and its role in parasitoid development and host regulation. Ann. Entomol. Soc. Am. 88: 800-808. Morales-Ramos, J. A., K. R. Summy and E. G. King. 1995b. Estimating parasitism by Catolaccus grandis (Hymenoptera: Pteromalidae) after inundative releases against the boll weevil (Coleoptera: Curculionidae). Environ. Entomol. 24: 1718-1725. Morales-Ramos, J. A., M. G. Rojas, and E. G. King. 1996. Significance of adult nutrition and oviposition experience on longevity and attainment of full fecundity of Catolaccus grandis (Hymenoptera: Pteromalidae). Ann. Entomol. Soc. Am. 89: 555-563. Morales-Ramos, J. A., M. G. Rojas, R. J. Coleman and E. G. King. 1998. Potential use of in vitro-reared Catolaccus grandis (Hymenoptera: Pteromalidae) for biological control of the boll weevil (Coleoptera: Curculionidae). J. Econ. Entomol. 91: 101-109. Quicke, D. L. J. 1997. Parasitic Wasps. Chapman and Hall, New York. Rojas, M. G., J. A. Morales-Ramos and E. G. King. 1996. In vitro rearing of the boll weevil (Coleoptera: Curculionidae) ectoparasitoid Catolaccus grandis (Hymenoptera: Pteromalidae) on meridic diets. J. Econ. Entomol. 89: 1095-1104. Rojas, M. G., J. A. Morales-Ramos, E. G. King, G. Saldaña and S. M. Greenberg. 1998. Use of a factitious host and supplemented adult diet to rear and induce oogenesis in Catolaccus grandis (Hymenoptera: Pteromalidae). Environ. Entomol. 27: 499-507. Rojas, M. G., J. A. Morales-Ramos, and E. G. King. 1999. Response of Catolaccus grandis (Hymenoptera: Pteromalidae) to its Natural Host After Ten Generations of Rearing on a Factitious Host (Callosobrucus maculatus) (Coleoptera: Bruchidae). Environ. Entomol. 28: 137-141. Summy, K. R., J. A. Morales-Ramos, E. G. King, S. Greenberg, and R. J. Coleman. 1994 a. Integration of boll weevil parasite augmentation into the short-season production system of the Lower Rio Grande Valley, pp. 953-957. In Proc. Beltwide Cotton Conferences 1994 Vol. 2. National Cotton Council of America, Memphis, TN. Summy, K. R., J. A. Morales-Ramos, E. G. King, R. J. Coleman and A. W. Scott. 1994b. Impact of mechanical cultivation on the searching efficiency of Catolaccus grandis, an exotic parasite of boll weevil. Southwest. Entomol. 19: 379-384. Summy, K. R., J. A. Morales-Ramos and E. G. King. 1995a. Suppression of boll weevil infestations on South Texas cotton by augmentative releases of the exotic parasite Catolaccus grandis (Hymenoptera: Pteromalidae). Biological Control 5: 523-529. Summy, K. R., J. A. Morales-Ramos, E. G. King, S. M. Greenberg, M. A. Rankin, L. G. Hansen and C. Moomaw. 1995b. Rationale and potential for augmentative biological control of boll weevil on follow-season cotton in southern Texas. Southwest. Entomol. 20: 483-491. Summy, K. R., S. M. Greenberg, J. A. Morales-Ramos, and E. G. King. 1997. Suppresion of boll weevil infestations (Coleoptera: Curculionidae) occurring on follow-season cotton in Southern Texas by augmentative releases of Catolaccus grandis (Hymenoptera: Pteromalidae). Biological Control 9: 209-215. Vargas-Camplis, J., R. J. Coleman, J. Gonzalez and L. Rodriguez Del B. 1997. Life table analysis of cotton boll weevil in the tropics of Tamaulipas Mexico after Catolaccus grandis releases, pp. 1194-1197. In Proceedings Belwide Cotton Conferences, 1997 Vol. 2. National Cotton Council of America, Memphis, TN. Waage, J. and D. Greathead. 1986. Insect Parasitoids. Academic Press, New York.
Back to Parasitoids Table of Contents
|
|
|||||||||||