|
||||||||||||
Back to Pathogens Table of Contents
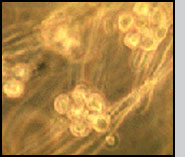
Lagenidium giganteum by James L. Kerwin, Mass Spectrometry Facility, Cornell University, Ithaca, NY 14853-2705 Lagenidium giganteum is a watermold that parasitizes the larval stage of mosquitoes. This microbial parasite belongs to a group of organisms that, although they look like fungi and have a "fungal lifestyle", nonetheless are related to diatoms and brown algae. The infective stage is a motile spore (see below) that goes on a search-and-destroy mission. The basis of its host specificity is selective recognition and attachment to its mosquito hosts. If a spore encounters, for instance, a water beetle, a dragon fly larva or a rice plant, it recognizes that a susceptible host has not been encountered. It will back off from that surface and swim on looking for a suitable host.
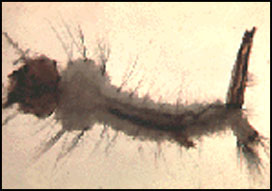
Appearance
(Note that infection by some other parasites, e.g. microsporidians, will also result in pale white larvae). In the absence of competing bacteria or protozoa, infected larvae will be completely filled with cells that under a microscope will appear translucent. These cells are often most easily seen either in the larval head capsule or the anal papilla (the breathing appartus at the tail end of the larva).
Habitat Although L. giganteum is not an obligate parasite, and can grow vegetatively (for example, on rotting vegetation or dead insects) in the absence of its hosts, it grows much faster and is easier to isolate from mosquito larvae. It can be found in freshwater habitats supporting mosquito populations. Isolation of the parasite from indigenous larval populations has been documented primarily in the southern United States, but it has also been found in California, Cuba, Colombia, and England. It will become dormant at temperatures below ca. 16°C or above 32°C. Moderate levels of salinity or organic load prevent it from sporulating, which is necessary for mosquito infection.
Pests Attacked The parasite will infect and kill most species of mosquito breeding in fresh water, from temperatures of 16-32°C. It will also infect the closely related dipteran Chaoborus astictopus, the Clear Lake gnat, and at very high concentrations, some species of daphnids. Infection of daphnids is not desireable, but this only occurs when levels of the parasite ca. 100 times greater than that recommended for operational control are reached.
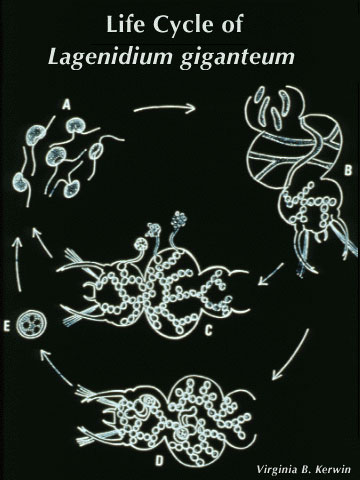
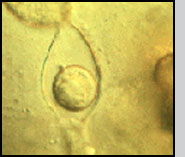
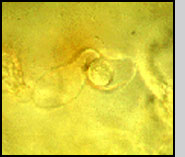
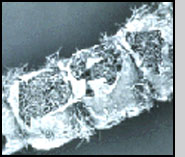
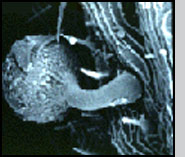
Life Cycle Infection of a larval host is initiated by motile biflagellate zoospores that selectively recognize chemical signals on the epicuticle (outer exoskeleton) of mosquitoes. After attaching (A) the zoospores inject themselves into the larva, and ramify throughout the body of the host (B). Depending upon the temperature and zoospore density, the larva dies of starvation within 1-4 days. At that time each individual cell can form an exit tube and release 10-50 asexual spores, which in turn seek out a new host (C). Alternately, two cells can fuse (D), ultimately resulting in the formation of a thick-walled dormant oospore (E). This sexual stage of L. giganteum can remain viable in a dehydrated state for at least 7 years. It is this spore that is responsible for multi-year recycling of the parasite even though a habitat may be dry for months or years before reflooding and colonization by mosquito larvae. Under appropriate environmental conditions, oospores will germinate, resulting in the production of infective biflagellate zoospores similar to those produced during asexual reproduction. Relative Effectiveness Operational levels of mosquito control have been obtained by ground or aerial application of L. giganteum at rates ranging from ca. 0.9 x 1010 to 5 x 1010 CFU's (colony forming units) per hectare. The application rate depends upon the susceptibility and developmental rate of the target species, and habitat characteristics (temperature, organic load and salinity). For instance, control of floodwater Aedes species in early fall, in which there is synchronous hatch of large numbers of eggs in relatively cold water, would require treatment at the higher rates. At the other extreme, for 3- to 4-month control of very susceptible mosquito species such as Culex tarsalis breeding in rice fields at very low densities, the lower rates can be used. Zoospores do not have a cell wall, so are much too fragile to be applied directly in a breeding habitat; therefore, either presporangia (mycelia, USEPA registration No. 56984-2), oospores (USEPA registration No. 56984-3), or a mixture of both (USEPA registration No. 56984-1) are applied. Sporulation then occurs in the treated site to initiate infection. The sexual stage of L. giganteum has many advantages in an operational control program including multi-year stability, resistance to desiccation and abrasion, and it's inherent slow-release characteristics. Unfortunately, fermentation yields of oospores remain 2 orders of magnitude below that of the less stable mycelial (asexual, presporangial) stage. Research is continuing on improving oospore yields, which would be much more useful in large scale operational mosquito control programs. L. giganteum is a facultative parasite, and has been grown in large fermentation tanks using inexpensive culture media. Multi-hectare aerial applications have been made in rice fields and in refuges for migratory birds, primarily in California, where both efficacy and safety to nontarget organisms has been documented. Because the mycelial cells are relatively rugged, the low volume spraying equipment used by many mosquito abatement agencies for application of insecticides is readily adapted to application of this parasite.
Pesticide Susceptibility Except for the dormant oospore stage, the parasite is very susceptible to organophosphates, carbamates, herbicides, fungicides, and other chemicals commonly used either by mosquito abatement districts, or applied to crops (such as rice and soybeans) that are associated with mosquito breeding. IC50 values for inhibition of mycelial growth commonly range from ca. 50-5,000 ppm. Fortunately, recommended application rates for many of these chemicals are below levels that are toxic to the parasite. Organic solvents such as toluene and oil commonly used as diluents of the active ingredients of pesticides are also toxic to L. giganteum.
Commercial Availability This parasite is registered with the U.S. Environmental Protection Agency and several states, including California and Florida, for use as an operational mosquito control agent. It is currently the only commercially available biological control agent (not including Bacillus thuringiensis var. israelensis, which is a microbial insecticide) for mosquitoes. Besides being host specific, L. giganteum has the ability to recycle for weeks, months, or even years in a given breeding habitat after a single application. Beginning in 1997 the asexual stage of this parasite will be commercially available as the LAGINEX® formulation. It can be obtained from AgraQuest, Inc.
References Brey, Paul T. and Remaudiere, G. (1985) Recognition and isolation of Lagenidium giganteum Couch. Bull. Soc. Vector Ecol. 10, 90-97. Domnas, Aristotle J. and Warner, Steven A. (1991) Biochemical activities of entomophagous fungi. Critical Reviews in Microbiology 18, 1-13. Kerwin, James L., Dritz, Deborah D. and Washino, Robert K. (1994) Pilot scale procution and application in wildlife ponds of Lagenidium giganteum (Oomycetes: Lagenidiales). J. Amer. Mosq. Control Assoc. 10, 451-455. Kerwin, James L. and Petersen, E.E. (1997) Fungi: Oomycetes and Chytridiomycetes. In Manual of Techniques in Insect Pathology (L. A. Lacey, ed.) pp. 251-268, Academic Press, New York. Lacey, Lawrence L. and Undeen, Al H. (1986) Microbial control of black flies and mosquitoes. Annu. Rev. Entomol. 31, 265-296. * Diagram reprinted from Manual of Techniques in Insect Pathology (L. A. Lacey, ed.), James L. Kerwin and Erin E. Petersen, "Fungi: Oomycetes and Chytridiomycetes," p. 252, 1997, by permission of the publisher Academic Press Limited London.
No endorsement of named or illustrated products is intended, nor is criticism implied of similar products that are not mentioned or illustrated.
Back to Pathogens Table of Contents
|
|
|||||||||||