|
||||||
Back to Pathogens Table of Contents
Bacillus thuringiensis, bio-pesticide By Zachary P. Cohen, Cornell University Last updated Feb 25, 2015
History Bacillus thuringiensis (Bt) is a bacterium that was first identified by S. Ishiwata in 1901 in Japanese silkworms presenting flacherie, or flaccid disease. It was later scientifically described and named by E. Berliner in Thüringen, Germany (Knowles 1994) . Berliner noted that the bacterium was producing insecticidal crystal (Cry) proteins, which were causing cell death in the digestive tract. These proteins have been used in insecticidal sprays and dusts to control pest insects since the 1930s. However, today they are most widely utilized as a transgene inserted into crop plants. These transgenic plants (Bt crops) are able to express the bacterial toxin-genes to defend themselves against herbivory. The use of these crops has been effective in controlling pest populations and increasing crop yield and quality. Bt crops were first commercialized in 1996 and, in 2013 187.5 million acres of Bt transgenic crops were planted worldwide (James et al. 2014).
Appearance/habitat Bt is a Gram-positive bacterium found in soil, plant, aquatic, and agricultural environments. During its vegetative state, the rod-shaped bacterium has the dimensions of 1.0-1.2 x 3.0-5.0 microns and occurs in long to short tangled chains. During sporulation, the ellipsoidal spore measures 0.5-1.2 x 1.3-2.5 microns and resides in the sporangium with parasporal crystalline toxins. The parasporal crystalline insecticidal proteins are as large or larger than the Bt spores (Lingren. 1984). Bt has a long and safe history of use to humans and the environment.
Applications The following are insecticidal formulations that can be applied topically in a private or commercial setting to control insect pests. There are many products currently registered by the EPA that contain Bt proteins and that target insect pests. Some of the products targeting caterpillars, including Dipel®, Biobit®, Condor®, Javelin®, CoStar ®, Crymax ® and Deliver®, are derived from toxins expressed in Bacillus thuringiensis var. Kurstaki, Btk. The formulations Agree® and XenTari® are based on the Bt strain Bacillus thuringiensis var. aizawai (Bta) that produces different proteins and has high efficacy against other caterpillars (Soberon et al. 2009). The Bt strain tenebrionis expresses proteins such as Cry3 toxins that kill coleopteran (beetle) grubs; these toxins are used in M-Trak®, Foil®, and Novodor®. Finally, mosquitocidal formulations derived from Bacillus thuringiensis israelensis are based on Cry4, Cry10, Cry11 toxins, and are publicly sold as Vectobac®, Teknar®, Bactimos®, Skeetal®, and Mosquito Attack® (Soberon et al. 2009). Since 1996 transgenic crops expressing Bt proteins have been commercially available in the US and other parts of the world. The presently commercial Bt crops in the US are corn and cotton. Bt soybeans are commercialized in some countries in South America and Bt eggplant was commercialized in Bangladesh in 2014. Many other Bt crops are under development for future agricultural use (James, 2014).
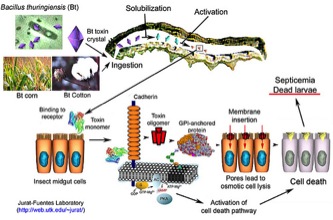
Mode of Action Unlike many synthetic insecticides, the insecticidal proteins produced by Bt strains are very specific for certain insect species. For example, some only affect caterpillars while others affect beetles, but none of them affect mammals. The many proteins produced are named and categorized based on their primary amino acid sequence and the type of insects they affect. Because of the narrow spectrum of activity against target insects, they are considered environmentally safe for birds, mammals, natural insect enemies that help suppress pest populations, and aquatic animals. For these reasons, the applications of these bio-insecticidal proteins are widespread and the topical formulations are used in organic farms. However, persistent use of topical solutions in an agricultural setting has led to cases of resistance in several species. Additionally, there are a few cases of insects having evolved resistance to transgenic plants expressing Bt proteins (Tabashnik et al. 2013). Crystalline Toxin Mode of Action Upon ingestion, the Bt protein is activated by alkaline (pH>7)-dependent proteins, which change the Bt protein into an active form within the insect’s digestive tract. This activation allows the protein to bind to receptors located on the intestinal lining. Once the protein binds, it forms holes in the gut that lead to cell death, septicemia, and insect mortality. The dead insect then supplies a nutrient- rich environment for the bacteria to propagate (Bravo et al. 2011). Changes in protein activation, diffusion, or receptor binding have all been associated with resistance in various insect pests. The first species to exhibit resistance to Bt toxins in an agricultural setting was the diamondback moth, Plutella xylostella (Syed, 1990). Populations in Malaysia and the Philippines that showed resistance were immune to the Btk derived formulation Dipel®, which expresses Cry1Aa, Cry1Ab, Cry1Ac, Cry2A and Cry2Ab (Iqbal et al., 1996). With increased use of Bt sprays, resistance to Bta and Btk formulations arose in Hawaii, Pennsylvania, Florida, and the Philippines (Shelton et al., 1993). The mechanisms of resistance in P. xylostella have been extensively investigated and were shown to have similar qualities between geographically isolated populations (Tabashnik et al. 1997). To categorize this phenomenon, Tabashnik devised a system to objectively differentiate a type of Bt resistance known as Mode 1 from other heritable forms of resistance.
Figure 3. Insecticidal protein mode of action. [http://web.utk.edu/~jurat/Btresearchtable.html] Mode 1 resistance is characterized by >500-fold resistance to at least one Cry1A protein via reduced binding at the midgut membrane, with little cross-resistance to other Bt proteins (Tabashnik et al. 1998). Diamondback moth strains from Hawaii, Pennsylvania, Florida, the Philippines, and South Carolina have developed resistance to Bt sprays and all exhibit characteristics of Mode 1 resistance (Tabashnik et al 1998). Non-Mode 1 resistance to Bt crops has arisen in three insect pest species of corn: Western corn rootworm, D. virgifera virgifera, fall armyworm, Spodoptera frugiperda, and maize stalk borer, Busseola fusca; and one to transgenic cotton: pink bollworm, Pectinophora gossypiella (Tabashnik et al 2013). Only resistance in one pest, S. frugiperda, in Puerto Rico has caused the crop to be taken off the market (Moar et al. 2008). However, strong resistance to Cry1F corn among populations of this species has recently caused crop loss in Brazil (Farias et al., 2014).
References Bravo, A., Likitvivatanavong, S., Gill, S. S., & Soberón, M. (2011). Bacillus thuringiensis: A story of a successful bioinsecticide. Insect Biochemistry and Molecular Biology, 41(7), 423–31. doi:10.1016/j.ibmb.2011.02.006
Back to Pathogens Table of Contents
|
|
|||||